Аномалии половых хромосом при нарушении репродуктивной функции у мужчин
|
(обзор литературы)
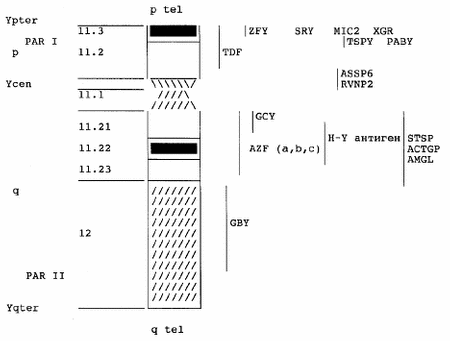
С.Г. Ворсанова, В.О. Шаронин, Л.Ф. Курило Московский НИИ педиатрии и детской хирургии Минздрава РФ; Медико-генетический научный центр РАМН, Москва У пациентов с генетическими нарушениями репродуктивной функции довольно часто встречаются синдромы, связанные с аномалиями в системе половых хромосом (гоносом) [18, 35, 70]. Исследования, проведенные у пациентов мужского и женского пола, свидетельствуют о том, что аномалии хромосом X и Y составляют около 2/3 всех хромосомных нарушений [19, 70, 98]. Частота синдромов, связанных с аномалиями гоносом, составляет 4,6 на 1000 новорожденных [6]. Известно, что около 2% мужчин страдают инфертильностью [55]. Среди мужчин с бесплодием и нарушением сперматогенеза у 5 — 15% обнаруживают хромосомные нарушения, аномалии гоносом составляют 75%, аутосом — 25% [13, 19, 29]. Частота врожденных нарушений полового развития у детей Москвы при рождении составляет 1 на 1550 [24]. При аномалиях гоносом отмечается отсутствие множественных пороков развития и умственной отсталости. При патологии гоносом лишь около 1% больных имеют задержку умственного развития [21]. При этом отставание выражено значительно меньше, чем при аномалиях аутосом [113]. Аномалии гоносом делят на количественные (моносомии и полисомии) и структурные (транслокации, делеции, дупликации, инсерции, инверсии), представленные полными и мозаичными формами [7—10, 119]. Наиболее распространенными являются синдромы Клайнфельтера (кариотип 47,ХХY; частота до 1,5 на 1000 новорожденных), дисомии Y (кариотип 47,XYY; частота 1 на 1000 новорожденных). Отмечено, что структурные перестройки хромосомы Y достаточно редки [18, 19, 90, 95, 114]. Причиной генетически обусловленного мужского бесплодия, связанного с нарушением сперматогенеза, является мутация в одном из локусов хромосомы Y [27,115]. Полагали, что функция хромосомы Y у млекопитающих ограничивается контролем за дифференцировкой пола [50]. Но в 1976 г. были представлены доказательства значения хромосомы Y человека для сперматогенеза [109]. Анализируя терминальную делецию хромосомы Y у 6 стерильных мужчин с нормальным фенотипом и азооспермией, авторы предположили существование генного комплекса мужской фертильности в дистальном участке эухроматиновой части длинного плеча хромосомы Y (Yq11). Этот локус назвали фактором азооспермии (AZF). Локализация AZF в районе Yq11 была подтверждена молекулярными исследованиями [115]. С 1976 г. начались интенсивные исследования хромосомы Y (и была построена её цитогенетическая, физическая и патологическая карта (см. рисунок).
В норме хромосома Y — одна из самых небольших по длине в кариотипе человека. Она выявляется ярким свечением дистальной части длинного плеча при окрашивании акрихинипритом, DAPI, а также С-методом. В 70-х годах изучали варианты длины окрашенного С-блока хромосомы Y. Отмечено, что при значительных колебаниях длины гетерохроматинового блока хромосомы Y обычно не наблюдается выраженных фенотипических отклонений, в то время как делеции или дупликации даже небольших сегментов других хромосом обычно приводят к тяжелым порокам развития. Было сделано предположение, что Q-окрашиваемый сегмент хромосомы Y генетически не активен и представляет собой компактную структуру конституционного гетерохроматина [23]. Это было подтверждено гибридизацией in situ c высокоповторяющимися последовательностями ДНК (сателлитная ДНК) [47]. Дистальный гетерохроматин занимает от 1/2 до 2/3 длинного плеча хромосомы Y. Хромосома Y делится на три участка: эухроматиновое короткое плечо Yp11, эухроматиновая проксимальная часть длинного плеча Yq11 и гетерохроматиновый дистальный участок Yq12 [64] (см. рисунок). Эухроматиновые районы невариабельны в размерах [34]. Эти данные справедливы для короткого плеча хромосомы Y [104]. Хромосома Y человека включает около 50 млн. нуклеотидных пар (н.п.), причем 60% ДНК находится в гетерохроматиновой области Yq12. При помощи гель-электрофореза в пульсирующем поле физическая протяженность короткого плеча оценена примерно в 13 млн. н.п. [83]. Следовательно, эухроматиновая часть длинного плеча Y (Yq11) не должна превышать 7 млн. н.п. На хромосоме Y локализовано более 100 генов и последовательностей ДНК, функции 16 из них определены [27]. Функции генов короткого плеча хромосомы Y (Yp), транскрибирующихся на разных этапах формирования органов репродуктивной системы и во время сперматогенеза, являются предметом изучения многих исследователей [27,115]. В этом участке с помощью молекулярных методов были обнаружены 2 гена, транскрибирующиеся в клетках яичка, названные ZFY (белок цинковых пальцев — zinc finger protein) и TSPY (тестисспецифический белок — testis specific protein). Мутации этих генов у стерильных мужчин до сих пор не описаны, роль их в сперматогенезе не ясна. Ген ZFY имеет консервативную структуру и экспрессируется во всех тканях человека, но участок размером 3000 н.п. активен только в яичках половозрелых мужчин. Он ответствен за дифференцировку пола, однако обнаружен также у пациентов с женским фенотипом и кариотипом 46,XY (синдром Сваера) [27, 86]. Ген TSPY транскрибируется во время сперматогенеза, однако его роль в формировании морфологических структур половых желез и семявыводящих канальцев не изучена. Возможно это специфический фактор, функционирующий в премейотических половых клетках мужчин [79]. На коротком плече хромосомы Y (Yp11) локализован ген, который получил название SRY (определяющий пол участок — sex determining region). Он имеет консервативную структуру, ответствен за развитие яичек. Его фрагмент в 35 тыс. н.п. представляет минимальную последовательность ДНК хромосомы Y, достаточную для формирования мужского фенотипа [22, 88]. Полагают идентичность этого локуса тестисдетерминирующему фактору (TDF) [56, 79]. В длинном плече хромосомы Y картированы гены, ответственные за репродуктивную функцию. Помимо AZF (фактора азооспермии), обнаружен ген, кодирующий специфический для мужчин мембранный клеточный белок — Н-Y антиген [39], который долго считали первичным фактором регуляции пола [120]. Н-Y антиген локализован в непосредственной близости к локусу AZF [36]. Очевидно, функция этого белка связана с формированием семенных канальцев. Имеются и другие гены длинного плеча хромосомы Y, не участвующие в формировании и функционировании половых органов. Ген GBY (гонадобластома—gonadoblastoma) рассматривается как онкоген, провоцирующий опухоли в дисгенетичных гонадах, развивающихся при мозаичных формах с кариотипом 45,Х/46,XY у индивидуумов с мужским и женским фенотипом [106]. Поэтому необходимо определение маркерных хромосом, что может быть достигнуто исключительно с помощью молекулярно-цитогенетических методов [116 — 118]. При нормальном кариотипе ген GBY проявляет активность перед сперматогенезом [86, 87]. Другой ген — GCY контролирует рост (growth control), локализован в проксимальной части Yq11 [30]. Его делеция или нарушение в нем последовательности ДНК обусловливает низкорослость. Cуществует мнение, что фактор, ответственный за рост, расположен в псевдоаутосомном регионе, в дистальной части Yp [59, 60]. Предполагают, что на хромосоме Y локализован не один, а несколько генов, контролирующих рост [38, 122]. Ген, контролирующий гаметогенез у мужчин, расположен в эухроматиновой части длинного плеча хромосомы Y-фактор азооспермии (AZF) [58]. Хотя AZF присутствует во всех клетках, кроме эритроцитов, он активен только в клетках Сертоли [115, 129]. В 1996 г. с помощью молекулярных методов была установлена неоднородная структура этого гена. Микроделеции, вызывающие нарушения репродуктивной функции у мужчин, были картированы в трех различных субрегионах Yq11. Один из субрегионов соответствующий локусу AZF, располагается в дистальной части Yq11, второй и третий картированы проксимальнее первого. У пациентов с микроделециями этих локусов выявлено нарушение сперматогенеза на различных стадиях, в зависимости от потери определенного участка AZF. Можно предположить наличие, как минимум, трех локусов в Yq11, контролирующих сперматогенез, причем каждый из них ответствен за разные стадии этого процесса. Последствием нарушения в каждой из указанных последовательностей является азооспермия или олигозооспермия тяжелой степени, поэтому эти локусы получили названия AZFa, AZFb и AZFc. Изучается их роль в процессе сперматогенеза [115]. Известно, что дистальные участки коротких плеч хромосом Х и Y человека подобны аутосомным [97], эти участки определяются как псевдоаутосомные районы (PAR I). Их физическая протяженность на хромосоме Y насчитывает менее 3 млн н.п. [94]. Известными гомологичными последовательностями хромосом Х и Y человека являются:1) дистальные участки коротких плеч хромосом Х и Y, соответствующие псевдоаутосомному региону (PAR I); 2) Х центромерные, Хр22 и Yq11 — 12 участки; 3) Хq13 и Yp12 участки; 4) Хq и Yq околотеломерные последовательности, иногда определяемые как псевдоаутосомный регион II (PAR II). Гомологичные последовательности расположены в коротких и длинных плечах хромосом Х и Y [27, 53, 91, 105]. Хромосомы Х и Y имеют от 10 до 14 высокогомологичных локусов [27, 85, 91, 92]. Такими гомологичными генами являются ZFX и ZFY, AMELX и AMELY (гены амелогенина), XG и XGPY (гены, определяющие группу крови) и некоторые другие. В настоящее время на гоносомах человека локализовано около 3 Mb гомологичных последовательностей. Показано, что на стадии зиготены профазы I мейоза хромосомы Х и Y соединяются участками своих коротких плеч. В этом районе половых хромосом наблюдают процесс кроссинговера и иногда аномальные Х — Y-обмены, способные привести к нарушению дифференцировки пола [14, 15, 59, 92]. В мейозе у мужчин частота рекомбинаций в 9,3 раза выше, чем у женщин [60]. На хромосоме Y человека расположены гены, контролирующие дифференцировку пола, формирование яичек и процесс сперматогенеза [68, 80]. Потеря или нарушение структуры этих генов, как правило, приводит к нарушению развития половой системы [22, 79, 92]. До появления метода дифференциального окрашивания хромосом определение структурных аномалий у хромосомы Y было невозможно. Выраженный полиморфизм длины хромосомы Y в нормальных мужских кариотипах не позволял сопоставить фенотипические аномалии выявленными нарушениями хромосомы Y. Как правило, аномальные хромосомы Y встречались в сочетании с клеточной линией 45,Х [71]. Поскольку имелось большое разнообразие фенотипов при мозаицизме Х/ХY (при аномальной хромосоме Y), включая аномалию пола, возник вопрос, определяется ли фенотипическое разнообразие нарушениями хромосомы Y или присутствием клеточной линии 45,Х. Различные типы дисгенезии гонад, в том числе интерсексуальные состояния, были выявлены у пациентов с кариотипом Х/ХY [46]. С появлением метода окрашивания акрихинипритом выяснилось, что нормальная по длине хромосома Y у стерильных мужчин с мозаичным кариотипом может полностью утрачивать гетерохроматиновый блок [40, 41]. Так как размер хромосомы Y был нормальным, она получила название “nl Y nf” (нефлюоресцирующая хромосома Y нормального размера). Позже Ynf хромосомы были определены как дицентрические изохромосомы с точками разрыва в Yq11 и с инактивированной второй центромерой [43, 106]. При немозаичных случаях кариотипа с Ynf, за редким исключением, проявляется мужской фенотип [54]. Возможно, это связано с дозовым эффектом гена SRY. Мозаичные кариотипы включают иногда более двух клеточных линий с различными Ynf хромосомами, что свидетельствует о нестабильности Ynf [31, 43]. Молекулярный анализ Ynf говорит о нарушении структуры гена AZF [54]. У таких пациентов наружные гениталии могут быть сформированными нормально или иметь неопределенное строение в зависимости от присутствия клеточной линии 45,Х. Обычно обнаруживается микроорхизм (гипоплазия яичек). У некоторых пациентов выявлены нарушения сперматогенеза на ранних стадиях мейоза [43]. Примечательно,что у стерильных мужчин с Ynf хромосомой не развивается гонадобластома, что является аргументом в пользу расположения гена GBY в Yq12 [73]. У больных обнаруживают общие клинические признаки с пациентами, имеющими моноцентрическую делетированную Yq хромосому. Впервые кариотип 46,X,Yq– был описан у мужчины с аспермией в 1973 г. [84]. Поскольку до этого были cообщения о нормальных фертильных мужчинах с таким же кариотипом, корреляция кариотипа и фенотипа была сомнительна. После выявления частичной делеции дистальной части эухроматинового района Yq у 6 мужчин с азооспермией установлено, что этот участок несет генетическую информацию, связанную с дифференцировкой мужских половых клеток [109]. Все пациенты имели нормальные наружные гениталии и гипопластичные яички. По гистопрепаратам биоптата яичка установлено отсутствие у одних половых клеток и клеток Сертоли, у других — выявлены только клетки Сертоли (при отсутствии половых клеток) [32, 57, 109]. При обнаружении крупных делеций Yq– в сочетании с азооспермией у мужчин наблюдается низкий рост [26, 67, 123]. Так, расположение фактора, контролирующего рост (GCY), было определено как проксимальное по отношению к AZF [30]. Менее распространенными структурными аномалиями являются кольцевые хромосомы Y. Образование такой хромосомы происходит при соединении точек разрыва в коротком и длинном плечах хромосомы Y. В большинстве случаев обнаруживается мозаицизм с клеточной линией 45,Х, описаны и полные формы 46,X,r(Y). У одного пациента с таким кариотипом был синдром Шерешевского — Тернера при мужском фенотипе [111]. Cтерильные мужчины с кольцевой хромосомой Y в кариотипе часто имеют нормальные наружные половые органы, но страдают азооспермией [42,46]. В ткани яичек стерильных мужчин с кольцевой хромосомой Y выявляют раннюю остановку сперматогенеза при полном отсутствии зрелых половых клеток, как при Ynf и Yq–. Кольцевая хромосома Y препятствует нормальной конъюгации гоносом в зиготене профазы I мейоза, что и вызывает нарушение развития сперматоцитов I и азооспермию. Особый интерес представляют Y/гоносомные транслокации. Транслокации между двумя хромосомами Y мало изучены и могут отличаться от Yq изохромосом размерами обоих плеч [128]. Часто встречается мозаицизм с клеточной линией 45,Х. Наружные половые органы у пациентов гипопластичны, в яичках выявляются только незрелые половые клетки до сперматоцитов I, указывающие на блок сперматогенеза в профазе I мейоза, что предполагает нарушение конъюгации Y/Y деривата с хромосомой Х во время зиготены [121]. В случаях транслокации участка хромосомы Y на хромосому Х пациенты чаще всего имеют мужской фенотип, характерны бесплодие, азооспермия. При этом наиболее распространены кариотипы 46,Y,der(X)t(X;Y)(p22.3q11) и 46,X,der(X)t(X;Y)(p22.3.q11) [53]. Цитогенетически не выявляемые Х;Y транслокации часто наблюдаются у стерильных мужчин с кариотипом 46,ХХ [16, 91]. Эта патология выявляется при флюоресцентной гибридизации in situ (FISH) c зондами, специфичными для определяющего пол участка короткого плеча хромосомы Y (ген SRY) [48, 75]. Мужчины с кариотипом 46,ХХ фенотипически сходны с больными с синдромом Клайнфельтера [16—18, 44], но отличаются нормальным ростом, размером зубов и отсутствием умственной отсталости. Синдром Ла Шапелля с кариотипом 46,ХХ у пациентов с мужским фенотипом встречается с частотой 1 на 20 000 — 24 000 новорожденных мальчиков. У этих пациентов отмечают азооспермию. Доказано, что у большинства пациентов материал хромосомы Y транслоцирован на короткие плечи одной из хромосом Х [52, 93]; длинное плечо хромосомы Y в таких случаях утрачивается, отсутствует и локус AZF, что объясняет азооспермию. Синдром Сваера — кариотип 46,ХY и женский фенотип. Частота этой патологии 1 на 100 000 новорожденных девочек. У больных линейные соединительнотканные тяжи (steak) вместо гонад (агенезия или дисгенезия гонад). Из-за отсутствия клеток Сертоли в них не вырабатывается антимюллеров гормон, чем и объясняется сохранение мюллеровых протоков; в плазме уровень ЛГ и ФСГ повышен. Женский фенотип и отсутствие тестикулярной ткани указывают на нарушения формирования половых органов по мужскому типу на начальных этапах [1, 17]. В отличие от мужчин с кариотипом XX, у женщин с кариотипом XY может быть выявлена делеция небольшого участка хромосомы Y. Описаны случаи XY-агонадизма у сестер, имевших двух нормальных братьев [65]. Таким образом, материал хромосомы Y, недостаточный для ХY- и избыточный для ХХ-кариотипа, крайне вариабелен, что приводит к различным нарушениям строения внутренних и наружных половых органов [49,79]. Известны случаи реверсии пола (женский фенотип, кариотип 46,ХY) за счет дупликации в участке короткого плеча хромосомы Х. Этот локус — DSS (Dosage Sensitive Sex reversal) расположен в Xp21 и составляет 160 kb. Возможно этот ген играет роль “посредника” при формировании половых органов по мужскому или женскому типу [130]. При сравнении гистологии яичек стерильных мужчин, имеющих в кариотипе Ynf, Yq– или r(Y) хромосому, с таковой у пациентов с Y-транслокациями, установлено у последних отсутствие дифференцированных половых клеток [114]. По-видимому, степень генетической активности хромосомы Х также влияет на развитие пола, так как инактивация хромосомы Х у мужчин с кариотипом 46,ХХ не случайна. Очевидно, нормальный процесс инактивации генов хромосомы Х может быть нарушен разрывом в Хр и присоединением участка короткого плеча хромосомы Y. Анализ полученных данных и сведения литературы [103] позволяют сделать три принципиально важных вывода: — ген, определяющий пол, транслоцированный на другую хромосому, может неправильно экспрессироваться; — некоторый ген, отличный от SRY, ZFY и других известных генов, ответственных за формирование пола, может оказывать частичное влияние на дифференцировку гонад; — при дисгенезии или агенезии и нормальном кариотипе гонад аномалии половых желез могут быть объяснены воздействием тератогена. Y/аутосомные транслокации — транслокации с участием хромосомы Y. В хромосоме Y очень много повторяющихся последовательностей ДНК, особенно Alu и Bkm, а также альфоидных повторов. Это может вызывать рекомбинацию с аналогичными последовательностями аутосом, участвующих в транслокации [85, 97, 110]. В большинстве семейных случаев Y/A транслокаций дистальная гетерохроматиновая часть хромосомы Y транслоцируется на короткое плечо акроцентрической хромосомы. Данный феномен может быть объяснен ассоциацией этих хромосом с комплексом ХY в стадии ранней пахитены. Если точки разрыва находятся в Yq12, мужчина фертилен без аномалий фенотипа. Y/A транслокация описана и для стерильных мужчин, это свидетельствует о вариабельности расположения точек разрыва в Yq12 и возможности их возникновения в дистальной или медиальной части Yq11, что приводит к нарушению функциональной структуры гена AZF. Менее распространены транслокации хромосомы Y с участием других аутосом (групп A — C, E, F). Последствиями этого (в зависимости от расположения точек разрыва и воссоединения) является азооспермия, обусловленная потерей генов, в частности — нарушением конъюгации половых хромосом в мейозе, а также аномальная инактивация хромосомы Х [74]. При делеции хромосомы Y потеря гетерохроматина Yq12 и незначительной части эухроматина совместима с нормальным развитием гениталий и дифференцировкой пола. Делеции, распространяющиеся на более проксимальные участки Yq11, часто вызывают азооспермию. Интерстициальные микроделеции в Yq11 выявляются с помощью молекулярно-генетических методов. Возможности метода FISH c малокопийными ДНК-последовательностями позволят выявлять даже незначительные нарушения набора хромосом человека. Это может стать новой вехой в диагностике хромосомных синдромов, в том числе и вызываемых структурными аберрациями с участием хромосомы Y. Значительную часть нарушений репродуктивной функции у мужчин составляют анеуплоидии гоносом. Наиболее частой их формой является синдром Клайнфельтера. Частота синдрома Клайнфельтера колеблется от 0,5 до 2 на 1000 новорожденных мальчиков (полные и мозаичные формы) [12, 20, 37]. Среди пациентов с легкой степенью умственной отсталости этот синдром выявляется у 1 — 2,5%; cреди бесплодных мужчин он выявляется у 10 — 15% [21]. Интенсивные исследования синдрома Клайнфельтера начались после 1956 г. [50]. У пациентов в метафазных клетках обнаружен кариотип 47,ХХY, а в интерфазных — хроматин Х. Новорожденные с синдромом Клайнфельтера не отличаются от здоровых мальчиков, лишь при исследовании мазков слизистой полости рта могут быть обнаружены Х-хроматинположительные ядра; иногда выявляют гипоплазию яичек. Основные клинические признаки проявляются в пубертатном возрасте. Характерный признак — евнухоидное телосложение с длинными ногами, узкими плечами и тазом, встречается у 50% больных. Иногда отмечается отложение жира на бедрах, груди и нижней части живота. В среднем рост больных соответствует таковому у здоровых мужчин. Часто встречаются неправильная форма черепа, непропорциональность длины конечностей, гинекомастия, обусловленная повышенной продукцией эстрогенов клетками Лейдига [11, 28]; как правило, отмечается выраженная гипоплазия внутренних половых органов. В биоптатах гонад отмечают значительное снижение числа половых клеток: отдельные сперматогонии обнаруживаются лишь в единичных канальцах. К пубертатному периоду наблюдается усиленная пролиферация интерстициальных клеток, обнаруживаются дегенерация и гиалинизация семенных канальцев. У пациентов отмечают деформации ушных раковин, алопецию, катаракту, прогнатизм, поперечную ладонную складку, радиоульнарный синостоз, сколиоз, неврологические нарушения, пороки сердца, но эти признаки не имеют диагностического значения [12]. Наружные половые органы, как правило, сформированы по мужскому типу; характерен микроорхизм, который является одним из важнейших клинических критериев синдрома. Объем эякулята редко достигает 1,5 мл; выявляется азооспермия. При обнаружении олигозооспермии целесообразно проведение молекулярно-цитогенетического исследования для выявления мозаичной формы синдрома и определения наличия и доли нормального клона клеток с кариотипом 46,XY [25, 80]. При мозаичной форме синдрома с преобладанием клона 46,ХY описаны нормальные фертильные мужчины, имеющие повышенную частоту специфических и неспецифических хромосомных аберраций в сперматозоидах [45]. Известны случаи сочетания кариотипа 47,ХXY с другими хромосомными аберрациями. При этом наблюдаются фенотипические проявления смешанного типа. Около 15% пациентов с синдромом Клайнфельтера страдают олигофренией, обычно в степени легкой дебильности, что тоже не может считаться постоянным диагностическим критерием синдрома. При более глубоком нарушении интеллекта отмечают аутизм, мнительность, склонность к алкоголизму, асоциальное поведение [20]. Увеличение числа хромосом Х в кариотипе ведет, как правило, к выраженной задержке психомоторного развития и более широкому спектру врожденных пороков и микроаномалий. Например, при кариотипе 49,XXXXY отмечается комплекс резко выраженных нарушений: пренатальная гипотрофия, задержка роста, лицевые аномалии (гипертелоризм, эпикант, уплощение спинки носа, косоглазие, близорукость); аномалии ушных раковин, прогнатизм, клинодактилия мизинцев, плоскостопие. Половые органы недоразвиты и видоизменены, у всех больных выявляются гипогонадизм и крипторхизм. В гонадах половых клеток не обнаруживается, интерстициальные клетки, как правило, гипоплазированы [20]. Больные с кариотипом 49,XXXXY страдают глубокой олигофренией: их IQ в среднем достигает 40, тогда как соответствующий показатель у пациентов с классическим типом синдрома Клайнфельтера c кариотипом 47,ХХY составляет более 80. В настоящее время известно более 100 случаев данной патологии, установлена материнская природа хромосом Х [78]. У больных с классической формой синдрома Клайнфельтера исследовали родительское происхождение дополнительной хромосомы Х. В 49% случаев лишняя хромосома Х наследовалась от отца, в 51% — от матери [72]. В группе пациентов с отцовским происхождением добавочной хромосомы отмечено значимое увеличение возраста отца. В случае материнского происхождения лишней хромосомы Х возраст матерей был выше в подгруппе с нерасхождением в первом мейотическом делении, нежели во втором. Клинических различий между больными в зависимости от происхождения дополнительной хромосомы Х не выявлено [72]. Другим типом анеуплоидии гоносом является полисомия хромосомы Y. За последние четверть века полисомия хромосомы Y обнаружена у нескольких десятков мужчин. Частота синдрома 0,75 — 1 на 1000 мужчин [95,128]. Характерными признаками этой хромосомной патологии являются асоциальное поведение и различные психологические нарушения, имеющиеся у 35% больных; среди мужчин с различными психическими отклонениями и асоциальным поведением частота синдрома колеблется от 0,45 до 15%. Более чем у 30% пациентов с кариотипом 47,ХYY наблюдается нарушение репродуктивной функции [51, 63, 66]. Клеточная линия 47,XYY в кариотипе больных с анеуплоидией хромосомы Y иногда сочетается с клонами 45,X, 46,XY, 47,XXY, 48,XXYY. Описаны случаи мозаицизма 45,X/49,XYYYY и 47,XYY/48,XYYY/49,XYYYY у пациентов с психическими отклонениями и нарушением репродуктивной функции [51, 108]. Описано несколько случаев обнаружения кариотипа 48,XYYY, из которых в половине выявлен мозаицизм с присутствием нормальной клеточной линии 46,XY. У таких больных отмечаются неонатальная асфиксия, умственная отсталость, ожирение. Часто они являются пациентами психиатрических клиник, для них характерны транссексуальность, агрессивность и периоды депрессии. Азооспермия у этих пациентов обусловлена атрофией семенных канальцев и полным отсутствием сперматогенеза [108]. Другим типом анеуплоидии гоносом у мужчин является кариотип 48,XXYY, впервые описанный в 1960 г. и названный “дважды мужчина” [82]. К настоящему времени описано несколько десятков таких пациентов; определены их клинические характеристики, которые во многом совпадают с таковыми при классическом синдроме Клайнфельтера. Рост у таких больных обычно выше среднего, для большинства характерны незначительные скелетные аномалии. В нескольких случаях отмечено агрессивное и асоциальное поведение [35,89]. Многие авторы относят к синдрому Клайнфельтера все разновидности анеуплоидий гоносом у пациентов с мужским фенотипом и с увеличением числа хромосом Х и Y, а также различные их мозаичные формы. Однако отличительные клинические признаки носителей таких анеуплоидий от классических случаев синдрома Клайнфельтера (47,XXY), ставят под сомнение подобную точку зрения [18]. Численные и структурные аномалии гоносом и хромосомы Y являются распространенными причинами нарушения репродуктивной функции мужчин. Изучение подобных хромосомных аномалий цитогенетическими и молекулярно-цитогенетическими методами позволит установить удельный вес различных типов гоносомных аберраций в соматических и половых клетках у взрослых пациентов с нарушением репродуктивной функции и у детей с различными формами нарушения полового развития [70,119,125]. Медико-генетическое консультирование и современные методы диагностики пациентов с нарушением репродуктивной функции хромосомного генеза позволяют в значительной степени изучить эту патологию и в отдельных случаях предупредить такие процессы, как малигнизация половых желез. Своевременное эффективное цитогенетическое и молекулярно-цитогенетическое обследование пациентов с нарушением полового развития позволяет также осуществлять правильную коррекцию пола и дальнейшее ведение этих больных. В заключение следует отметить возрастающее значение новых методов в диагностике нарушений репродуктивной функции, обусловленных в первую очередь патологией хромосом. Методы молекулярно-цитогенетической идентификации хромосомных нарушений, основанные на использовании особого типа клонированных фрагментов ДНК человека — хромосомоспецифичных ДНК-проб, позволяют определить особенности хромосомного набора соматических и половых клеток без прямого цитогенетического анализа, при помощи гибридизации нуклеиновых кислот в условиях in situ [2—5, 119]. Используя молекулярно-цитогенетический метод, можно выявлять и анализировать нормальные или аномальные гомологичные хромосомы в клетках с различной степенью дифференцировки и практически на всех стадиях клеточного цикла, в том числе и в интерфазе, что особенно важно для обнаружения анеуплоидий в половых клетках человека [9, 100, 101, 124]. FISH-анализ в настоящее время становится одним из ведущих методов определения численных хромосомных нарушений в сперматозоидах и незрелых половых клетках человека [61, 76, 77, 112, 124]. C целью повышения эффективности выявления аномалий хромосом в интерфазных клетках, особенно таких, как высококонденсированные сперматозоиды, предложены новые модификации протокола FISH, в том числе для двух- и трехцветовой флюоресцентной гибридизации in situ, а также метода праймированного мечения in situ [33, 62, 81, 96, 107, 125, 127]. Другим, более универсальным и простым подходом, является метод быстрой FISH, позволяющий получить результаты в течение 5—15 мин [99, 124, 125]. В настоящее время проводятся интенсивные исследования по молекулярно-цитогенетическому картированию центромерных, теломерных и сайт-специфичных РАС-клонов, в том числе маркирующих определенные участки хромосомы Y [102,126]. Эти ДНК-пробы в перспективе могут быть использованы для диагностики как численных, так и структурных микроаномалий, затрагивающих хромосому Y, что позволит устанавливать корреляцию между нарушениями тонкой структуры хромосомы Y и фенотипическими последствиями таких аномалий. Достижения современной науки открывают принципиально новые возможности в разработке и усовершенствовании методических подходов к изучению репродуктивной функции у мужчин, в целом, и различных форм аномалий гоносом и аутосом, в частности, а также внедрить их в практику здравоохранения. Литература 1. Беникова Е.А., Бужиевская Т.И., Сильванская Е.М. Генетика эндокринных заболеваний. Киев, Наукова думка 1993;400. 2. Ворсанова С.Г., Юров Ю.Б. Молекулярно-цитогенетическая диагностика хромосомных синдромов. Использование достижений генетики в охране здоровья матери и ребенка (опыт стран — членов СЭВ).1987;88 — 94. 3. Ворсанова С.Г., Казанцева Л.З., Демидова И.А., Дерягин Г.В. Цитогенетическая диагностика у супружеских пар с отягощенным акушерским анамнезом. Вопросы охраны материнства и детства. 1989;34:6:52 — 56. 4. Ворсанова С.Г., Юров Ю.Б., Александров И.А. и др. Молекулярно-цитогенетическая диагностика наследственных болезней, связанных с различными аномалиями хромосом Х. Педиатрия. 1989;1:76 — 80. 5. Ворсанова С.Г., Юров Ю.Б., Соловьев И.В. Картирование генов и молекулярная диагностика наследственных болезней. Медицинская генетика (экспериментальная информация), 1989;11:1 — 16. 6. Ворсанова С.Г., Казанцева Л.З., Демидова И.А., Юров Ю.Б. Использование методов молекулярно-цитогенетической диагностики в практике генетического консультирования. Информационное письмо, 1990. 7. Ворсанова С.Г. Хромосомные аномалии у детей с недифференцированной олигофренией и супружеских пар с отягощенным акушерским анамнезом по данным молекулярно-цитогенетических исследований: Автореф. дисс. ј док. биол. наук. Киев 1991;52. 8. Ворсанова С.Г., Казанцева Л.З., Юров Ю.Б. и др. Использование молекулярно-цитогенетических методов диагностики в генетической клинике. Вопросы охраны материнства и детства. 1991;11:71 — 76. 9. Ворсанова С.Г., Юров Ю.Б., Дерягин Г.В. и др. Идентификация хромосом Х в интерфазных ядрах с помощью гибридизации нуклеиновых кислот in situ для выявления анеуплоидий. Бюлл экспер биол мед 1991;10:72 — 77. 10. Ворсанова С.Г., Казанцева Л.З., Берешева А.К., Демидова И.А. Результаты молекулярно-цитогенетической диагностики супружеских пар с нарушением репродуктивной функции при медико-генетическом консультировании. Молекулярная диагностика наследственных болезней и медико-генетическое консультирование. Сб-к научных трудов. М. МОНИКИ, 1995;Т.1:124 — 133. 11. Давиденкова Е.Ф., Верлинская Д.К.,Тысячник С.Ф. Клинические синдромы при аномалиях половых хромосом. Л. Медицина, 1973;198. 12. Козлова С.И., Демикова Н.С., Семанова Е., Блинникова О.Е. Наследственные синдромы и медико-генетическое консультирование. М. Практика, 1996;305. 13. Курило Л.Ф. Возможности цитогенетического исследования мейоза при мужском бесплодии. Цитол и генет 1989;23:2:63 — 70. 14. Курило Л.Ф., Любашевская И.А., Дубинская В.П., Гаева Т.Н. Количественный кариологический анализ состава незрелых половых клеток из эякулята. Урология и нефрология 1993;5:45 — 47. 15. Курило Л.Ф., Дубинская В.П., Остроумова Т.В. и др. Анализ патологии сперматогенеза различной этиологии по эякуляту. Пробл репрод 1995;3:33 — 38. 16. Курило Л.Ф., Козлов Г.И., Федотов В.П. и др. Клинико-цитогенетическое обследование пациента с кариотипом 46,ХХ, азооспермией и бесплодием. Пробл эндокринол 1994;40:12:50 — 51. 17. Курило Л.Ф. Некоторые этапы дифференцировки пола, развития половых клеток и органов половой системы. Пробл репрод 1996;2:62 — 70. 18. Курило Л.Ф. Генетически обусловленные нарушения мужской репродуктивной системы. Сб. Сексология и андрология. Киев 1996;28 — 46. 19. Курило Л.Ф., Шилейко Л.В., Мхитарова Е.В. и др. Структура наследственной патологии половой системы при обследовании пациентов с нарушением репродукции.Тез. конф.”Инвалидиз. наследственные заболевания”, МГНЦ РАМН, Москва, XI 1997. 20. Лазюк Г.И. Тератология человека. М. Медицина, 1991;434. 21. Маринчева Г.С., Гаврилов В.И. Умственная отсталость при наследственных болезнях. М. Медицина, 1988;255. 22. Осипова Г.Р. Исследование гена SRY при некоторых нарушениях детерминации пола (XY “чистой” дисгенезии гонад, синдроме Шерешевского—Тернера, ХХ-инверсии пола). Автореф. дисс. ј канд. мед. наук. М 1997;24. 23. Прокофьева-Бельговская А.А. Гетерохроматиновые районы хромосом. М Наука, 1986;430. 24. Тарская Л.А. Клинико-цитогенетический анализ аномалий полового развития (синдром Шерешевского-Тернера и смешанная дисгенезия яичек). Автореф. дисс. ... канд. мед. наук. М 1996;26. 25. Юров Ю.Б., Ворсанова С.Г., Бытенская Г.А. и др. Молекулярно-интерфазная пост- и пренатальная диагностика синдромов, связанных с аномалиями половых хромосом. Сб-к научных трудов. М. МОНИКИ, 1995;Т.1.:141 — 150. 26. Alvesalo L., Chapelle A. de la. Tooth size in two males with deletions of the long arm of the Y chromosome. Ann Hum Genet 1981;54:49 — 54. 27. Affara N., Bishop C., Brown W. et al. Report of the international workshop on Y chromosome mapping 1995. Cytogenet Cell Genet 1996;73:33 — 76. 28. Bandmann H.-J., Breit R., Perwein E. Klinefelter’s syndrome. Springer Verlay, 1984. Berlin Heidelbery New York Tokyo. 29. Barakat A.I., Seikaly M.G., Kaloustian V.M. Urogenital abnormalities in genetic diseases. J of Urology 1986;136:4:778 — 785. 30. Barbaux S., Vilain E., Delafontain D. et al. Deletions of the long arm of the Y chromosome define regions involved in male growth, spermatogenesis and Turner’s phenotype. European society of human genetics. 26th annual meeting. Paris, France, 1994;170. 31. Batsone P.J., Faed M.J.W., Jung R.T., Gosden J. 45,X/46,Xdic(Y) mosaicism in phenotypic male. Arch Dis Child 1991;66:252 — 253. 32. Beverstock G.C., Macfarlane J.D., Veenema H. et al. Y chromosome specific probes identify breakpoint in a 45,X/46,X, del(Y)(pter-q11.1:) karyotype of an infertile male. J Med Genet 1989;26:330 — 342. 33. Bischoff F.Z., Nguyen D.D., Burt K.J., Shaffer L.G. Estimates of aneuploidy using multicolor in situ hybridization on human sperm. Cytogenet Cell Genet 1994;66:237 — 243. 34. Bobrow M., Pearson P.L., Pike M.C., El-Alfi O.S. Length variation in the quinacrine-binding segment of human Y chromosome of different sizes.Cytogenetics 1971;10:190 — 198. 35. Borgaonkar D.S. Cromosomal variation in man. A catalog of chromosomal variants and anomalies. Alan R.Liss, Inc., NY, 5th edition, 1989. 36. Bourgoyne P. Y chromosome function in mammalian development. Developmental biology 1991;1:1 — 29. 37. Brothman R. Cytogenetic basis of human disease. Cellular and molecular pathogenesis, Liooincott-Raven Publishers, Philadelphia, 1996;445 — 460. 38. Buhler E.M. Clinical and cytologic impact of Y chromosome abnormalities. The Y chromosome. Ed. Sandberg A.A. Ptb.-New York, Alan R.Liss, 1985;61 — 93. 39. Cantrell M.A., Simpson E., Bicknell J.N. et al. Absence of H — Y anigene expression in a patient with deletion of the long arm of the Y chromosome and isolation of a new DNA probes for region 6 of the Y chromosome. Am J Hum Genet 1989;45:134. 40. Caspersson T., Faber S., Foley G.E. et al.Chemical differentiation along metaphase chromosomes. Exp Cell Res 1968;49:219 — 222. 41. Caspersson T., Zech L., Johansson J. et al. Translocations сausing non-fluorescent Y chromosomes in X0/XY mosaics. Hereditas.,1971;68:281 — 304. 42. Chandley A.C. Infertility and chromosomal abnormality. Oxford Rev Reprod Biol 1984;6:1 — 46. 43. Chandley A., Ambros P., McBeath S. et al. Short arm dicentric Y chromosome with associated statural defects in a sterile man. Hum Genet 1986;73:350 — 353. 44. Chapelle A. de la.The etiology of maleness in XX men. Hum Genet 1981;58:105 — 116. 45. Cozzi J., Chevret E., Rousseaux S. et al. Achievment of meiosis in XXY germ cells: study of 543 sperm karyotypes from an XY/XXY mosaic patient. Hum Genet 1994;93:32 — 34. 46. Daniel A. Y isochromosomes and rings. In: A.A.Sandberg (ed) The Y chromosome, Part B:Clinical aspects of Y chromosome abnormalities. Alan R. Liss, Inc., NY., 1985;105 — 135. 47. Evans H.J., Gosden J.R., Mitchell A.R., Buckland R.A. Location of human satellite DNAs on the Y chromosome. Nature 1974;251:346 — 347. 48. Farah S.B., Garmes H.M., Cavalacanti D.P. et al. Use of Y-chromosome-specific DNA probes to evaluate an XX male. Brazilian J Med Biol Res 1991;24:149 — 156. 49. Ferguson-Smith M.A., Affara N.A., Maginis R.E. Ordering of Y-specific sequences by deletion mapping and analysis of X-Y interchanges males and females. Development, 1991;149 — 156. 50. Ford C.E., Hamerton J.L. The chromosomes of man. Nature 1956;1020 — 1023. 51. Fryns J.P., Kleczkowska A., Kubien E., van den Berghe H. XYY syndrome and other Y chromosome polysomies. Mental status and psychological functioning. Genet Counsel 1995;6:3:197 — 206. 52. Fuse H., Satomi S., Katayama T. et al. DNA hybridization study using Y-specific probes in an XX-male. Andrologia 1991;23:237 — 239. 53. Gabriel-Robez O., Rumpler Y., Ratomponirina C. et al. Deletion of the pseudoautosomal region and lack of sex chromosome pairing at pachytene in two infertile men carrying an X;Y translocation. Cytogenet Cell Genet 1990;54:38 — 42. 54. Genuardi M., Bardoni B., Floridia G. et al. Dicentric chromosome Y associated with Leydig cell agenesis ans sex reversal. Clin Genet 1995;47:38 — 41. 55. Goncalves J., Rocha T., Vale F. et al. Delineation of genetic basis of azoospermia. Eur J Hum Genet 1996;4S1:21. 56. Harley N.R., Jackson D.I., Hextall P. et al. DNA binding activity of recombinant SRY from normal males and XY females. Science, 1992;255:453 — 455. 57. Hartung M., Devictor M., Codaccioni J.L., Stahl A. Yq deletion and failure of spermatogenesis. Ann Genet 1988;31:21 — 26. 58. Henegariu O., Hirschmann P., Kilian K. et al. Rapid screening of the Y chromosome in idiopathic sterile men, diagnostic for deletions in AZF, a genetic Y-factor expressed during spermatogenesis. Andrologia 1994;26:97 — 106. 59. Henke A., Wapenaar M., van Ommen G.L. et al. Deletions within the pseudoautosomal region help map three new markers and indicate a possible role of this region in linear growth. Am J Hum Genet 1991;49:811 — 819. 60. Henke A., Fischer C., Rappold G. Genetic map of the human pseudoautosomal region. European society of human genetics. 25th annual meeting. Barcelona, Spain, 1993;92. 61. Holmes J.M., Martin R.H. Aneuploidy detection in human sperm nuclei using fluorescence in situ hybridization. Hum Genet 1993;94:443 — 453. 62. van Hummelen P., Lowe X.R., Wyrobek A.J. Simaltaneous detection of structural and numerical chromosome abnormalities in sperm of healthy men by multicolor fluorescence in situ hybridization. Hum Genet 1996;98:608 — 615. 63. Ion R., Telvi L., Feingold J. et al. Y chromosome abnormalities associated with mental retardation and dismorphia. European society of human genetics. 26th annual meeting. Paris, France, 1994;171. 64. ISCN 1995. An international systeme for human cytogenetic nomenclature, Mitelman F. (ed); S.Karger,Basel, 1995;5 — 115. 65. Kennerknecht I., von Saurma P., Brenner R. et al. XY-agonadism in two sisters with multiple extragenital malformations. A new autosomal recessive syndrome. European society of human genetics. 26th annual meeting. Paris, France, 1994;66. 66. Kleczkowska A., Fryns J., van den Berghe H. X-chromosome polysomy in male: the Leuven experience 1966—1987. Hum Genet 1988;80:1:16 — 22. 67. Kosztolanyi G.,Trixler M. Yq deletion with short stature, abnormal male development, and schizoid character disorder. J Med Genet 1983;20:39 — 394. 68. Kucheria K., Mc Elreavey K., Fellous M. The present status of our understanding of human sex determination. Cytogenet Cell Genet 1997;77:113. 69. Kurilo L., Schapoval N., Dubinskaya V. et al. Mitotic and meiotic chromosome studies in 140 infertile males with spermatogenesis disturbances. Abstr.Int.ESHG, 26th ann.meet., Paris, P.126. 70. Kurilo L., Mchitarova E., Schilejko L. et al.Chromosomal anomalies in patients with reproductive failures. Abst. ESMG— 28th An Meet IV-1996, London. 71. Lin C.C., Meyne J., Sasi R. et al. Determining the origin and the structural aberration of small marker chromosomes in two cases of 45,X/46,X,+mar by use of chromosome-specific DNA probes. Am J Med Genet 1990;37:71 — 78. 72. Lorda-Sanchez I., Binkert F., Maechler M. et al. Reduced recombination and parental age effect in Klinfelter syndrome. Hum Genet 1992;5:524 — 530. 73. Lukusa T., Fryns J.P., van den Berghe H. Gonadoblastoma and Y-chromosome fluorescence.Clin Genet 1986;29:311 — 316. 74. Maraschio P.,Tupler R., Dainotti E. et al. Molecular analysis of a Y/1 translocationin in an azoospermic male. European society of human genetics. 25th annual meeting. Barcelona, Spain, 1993;109. 75. Margarit E., Ballesta F., Carrio A. et al. Molecular characterization of Y sequences in XX males and localization of translocations by FISH. Cytogenet Cell Genet 1997;77:115. 76. Martin R.H., Balkan W., Burns K. The chromosome constitution of 1000 human spermatozoa.Hum Genet 1993;63:305 — 330. 77. Martin R.H., Spriggs E., Ko E., Rademaker A.W. The relationship between parental age, sex ratios, and aneuploidy frequencies in human sperm, as assesed by multicolor FISH. Am J Hum Genet 1995;57:1395 — 1399. 78. Marton V., Nystad T., Mokleby E. et al. Origin of the X chromosomes in a patient with the 49,XXXXY syndrome determined by polymorphic CA repeat sequences. European society of human genetics. 25th annual meeting. Barcelona, Spain, 1993;102. 79. McElreavey K.D., Vilain E, Abbas N. et al. A regulatory cascade hypotesis for mammalian sex determination:SRY represses a negative regulator of male development. Proc Natl Acad Sci USA, 1993;90:3368 — 3372. 80. Micic M., Nikolis J., Micic S., Diklic V. Y chromosome anomalies and male fertility. Hellenic Association of medical geneticists and university of Thessaloniki. 1st Balkan meeting on human genetics. Greece, 1994;91. 81. Miharu N., Best R.G., Young S.R. Numerical chromosome abnormalities in spermatozoa of fertile and infertile men detected by fluorescence in situ hybridization. Hum Genet 1994;93: 502 — 506. 82. Muldal S., Ockey C.H. The “double male”: a new chromosome constitution in Klinfelter’s syndrome. Lancet 1960;ii:492 — 493. 83. Muller U., Lalande M. A phisical map of the human chromosome Y shot arm.Genomics 1990;7:517 — 523. 84. Neu R.L., Barlow M.J., Gardner L.I. A 46,XYq– male with aspermia. Fertil Steril 1973;24:811 — 813. 85. Ogata T., Tyler-Smith C., Purvis-Smith S., Turner G. Chromosomal localization of gene(s) for Turner stigmata on Yp. J Med Genet 1993;30:918 — 922. 86. Page D.C., Harper M.E., Love J., Botstein D. Occurence of a transposition from the X-chromosome long arm to the Y-chromosome short arm during human evolution. Nature 1984;311:119 — 123. 87. Page D.C. Hypotesis: a Y chromosomal gene causes gonadoblastoma in disgenetic gonads. Development 1987;101:151 — 155. 88. Palmer M.S., Sinclair A.H., Berta P. et al. Genetic evidence that ZFY is not testis-determining factor. Nature 1989;342:937 — 939. 89. Parker C.E., Mavalwala J., Melnyk J., Fish C.H. The 48,XXYY syndrome. Am J Med 1970;48:777 — 781. 90. Peterlin B., Kunej T., Zorn B. et al. Sterility associated with Y chromosome abnormalities. Cytogenet Cell Genet 1997;77:83. 91. Petit C.,Chapelle A.de la., Levillers J. et al. An abnormal terminal X-Y interchange accounts for most but not all cases of human XX maleness. Cell 1987;49:595 — 602. 92. Petit C., Levillers J., Rouyer F. et al. Isolation of Sequences from Xp22.3 and deletion mapping using sex chromosomes rearrangements from human X-Y interchange sex reversal. Genomics 1990;6:651 — 658. 93. Pinto C., Magalhaes R., Pinto Carvalho A.P. XX male — molecular diagnosis resources with Y probes. Eur J Hum Genet 1996;4S1:136. 94. Rappold G., Lehrach H. A long range restriction map of the pseudoautosomal region by partial digest PFGE analysis from the telomere. Nucl Asids Res 1988;16:5361 — 5377. 95. Rothwell N.Y. Understanding genetics: sex chromosome anomalies in humans, 3rd edn. NY:Oxford university press,1983;113 — 118. 96. Rousseaux S.,Chevret E. In vitro decondensation of human spermatozoa for fluorescence in situ hybridization. Hum Reprod 1995;10:2209 — 2213. 97. Rouyer F., Simmler M.-C., Johnsson C. et al. A gradient of sex linkage in the pseudoautosomal region of the human sex chromosomes. Nature 1986;319:291 — 295. 98. Shirley G., Tierney S., Ushaho J. The edinburgh study of growth and development of children with sex chromosome abnormalities. Birth Defects: Original article series., 1982;18:4:41 — 60. 99. Soloviev I.V., Yurov Y.B., Vorsanova S.G., Malet P. Microwave activation of fluorescence in situ hybridization: a novel method for rapid chromosome detection and analysis. Focus 1994;16:4:115 — 116. 100. Soloviev I.V., Yurov Y.B., Vorsanova S.G. Prenatal diagnosis of trisomy 21 using interphase fluorescence in situ hybridization of postreplicated cells with site-specific cosmid contig probes. Prenatal Diagn. 1995;15:237 — 248. 101. Soloviev I.V., Fayet F., Malet P. et al. Fluorescence in situ hybridization with a new approach for identification of trisomy 21 in interphases amniocytes using cosmid probes. Cytogenet Cell Genet 1995;69:119 — 120. 102. Soloviev I.V., Yurov Y.B., Ioannou I. et al. Identification and molecular-cytogenetic characterization of large subset of human plasmids, cosmids, PAC and YAC clones: the search of DNA probes for pre- and postnatal diagnosis. Cesko-Slovenska pediatrie 1997;7:529 — 538. 103. Sorgo W., Gortner L., Bartmann P. et al. Gonadal agenesis in 46,XY female with multiple malformations and positive testing for the sex-determination region of the Y chromosome. Horm Res 1991;35:124 — 131. 104. Soudek D., Langmuir V., Stowart D.J. Variation in the nonfluorescent segment of long Y chromosome. Hum Genet 1973;18:285 — 290. 105. Speed R.M., Chandley A.C. Prophase of meiosis in human spermatocytes analysed by EM microspreading in infertile men and their controles and comparisons with human oocytes. Hum Genet 1990;84:547 — 554. 106. Speleman F., Van der Auwera B., Mangelschots K. et al. Identification and characterization of normal length nonfluorescent Y chromosomes: cytogenetic analysis, Southern hybrydization, and nonisotopic in situ hybridizatin. Hum Genet 1990;85:569 — 575. 107. Spriggs E.L., Rademaker A.W., Martin R.H. Aneuploidy in human sperm: results of two- and three-color fluorescent in situ hybridization using centromeric probes for chromosomes 1,12,18,X, and Y. Cytogenet Cell Genet 1995;71:47 — 53. 108. Teyssier M., Pousset G. 46,XY/48,XYYY mosaicism case report and review of the literature. Genet Couns 1994;5:4:357 — 361. 109. Tiepolo L., Zuffardi O. Localization of factors controlling spermatogenesis in the nonfluorescent portion of human Y chromosome long arm. Hum Genet 1976; 34:119 — 124. 110. Tyler-Smith C., Taylor L. Structure of a hypervariable tandemly repeated DNA sequence on the short arm of the human Y chromosome. J Mol Biol 1988;203:837 — 848. 111. Tzancheva M., Tyler-Smith C., Kumanov P., Kaneva R. Turner’s phenotype in a male with ring Y chromosome. Eur J Hum Genet 1996;4:S1:149. 112. Vidal F., Egozcue J. The cytogenetics of human gamets. Cytogenet Cell Genet 1997;77:48. 113. Vogel E., Motulsky A.G. Human Genetic. NY., Springer-Verlag, 1986. 114. Vogt P. Y chromosome function in spermatogenesis. In: Spermatogenesis-Fertilization-Contraception. Molecular, cellular and endocrine events in male reproduction. Springer Verlag, NY, 1992;4:226 — 257. 115. Vogt P.H., Edelmann A., Kirsch S. et al. Human Y chromosome azoospermia factors (AZF) mapped to different subregions in Yq11. Hum Molec Genet 1996;5:7:933 — 943. 116. Vorsanova S.G., Yurov Yu.B., Passarge I. et al. Identification of marker chromosomes by in situ hybridization technique using alpha and “classical” satellite DNA probes with relative chromosomal specificity. Цитол и генет 1994;28:3:67 — 70. 117. Vorsanova S.G., Yurov Yu.B., Soloviev I.V. et al. Rapid identification of marker chromosomes by in situ hybridization under different stringency conditions. An Cell Patol 1994;7: 251 — 258. 118. Vorsanova S.G., Demidova I.A., Soloviev I.V. et al. Pre- and postnatal diagnosis of additional marker chromosomes. Cesko-Slovatska pediatrie 1997;7:544 — 550. 119. Vorsanova S.G., Yurov Y.B., Soloviev I.V. et al. Molecular-cytogenetic diagnosis of chromosomal anomalies in genetic counseling. Cesko-Slovatska pediatrie 1997;7:538 — 544. 120. Wachtel S.S. H — Y antigene and the biology of sex determination. Grune & Stratton, New York, 1983. 121. Wahlstrom J. Y/Y translocations and their cytologic and clinical manifistation. In: Sandberg A.A. (ed) The Y chromosome, Part B: Clinical aspects of Y chromosome abnormalities. (Alan R.Liss,Inc., NY)., 1985;207 — 212. 122. Weissenbach F., Goodfellow P. Report of the Comittee on the genetic constitution of the Y chromosome. Cytogenet Cell Genet 1990; 55:1 — 4:314 — 320. 123. Yunis E., Garcia-Conti L.,Torres de Caballero O.M., Giraldo A. Yq deletion, aspermia, and short stature. Hum Genet 1977;39:117 — 122. 124. Yurov Y.B., Saias M.J., Vorsanova S.G. et al. Rapid chromosomal analysis of germ-line cells by FISH: an investigation of infertile male with large-head spermatozoa. Molec Hum Reprod 1996;2:9:665 — 668. 125. Yurov Y.B., Soloviev I.V., Vorsanova S.G. et al. High resolution multicolor fluorescence in situ hybridization using cyanine and fluorescein dyes: ultra rapid chromosome detection by directly fluorescently-labeled alphoid DNA probes. Hum Genet 1996;97:390 — 398. 126. Yurov Y.B., Soloviev I.V., Vorsanova S.G. et al. DNA probes for pre- and postnatal diagnosis of chromosomal anomalies: a collection for FISH analysis. Cesko-Slovatska pediatrie 1997;7:550 — 554. 127. Zalensky A.O., Allen M.J., Kobayashi A. et al. Well-defined genome architecture in the human sperm nucleus. Chromosoma 1995;103:577—590. 128. Zelante L., Dallapicola B., Calvano S. et al. Two mosaic-YY males carring asymmetric Y chromosomes. Clin Genet 1997;51:65 — 68. 129. Zenzes M.T., Wolf U., Engel W. Organization in vitro of ovarian cells in testicular structures. Hum Genet 1978;44:333 — 338. 130. Zuffardi O.,Camerino G., Arrigo G. et al. Mechanisms of Y chromosome rearrangements and related phenotypes. Cytogenet Cell Genet 1997;77:40. |