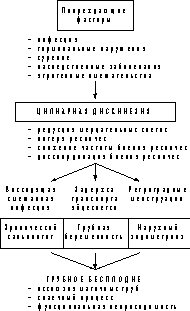
Цилиарная дискинезия в патогенезе трубного бесплодия.
|
Цилиарный транспорт в маточных трубах
Основная репродуктивная задача маточных труб - обеспечение капацитации сперматозоидов, оплодотворения яйцеклетки, жизнедеятельности эмбриона на ранних стадиях развития и осуществление переноса его в полость матки. Этому способствует слаженная работа рецепторного аппарата эпителия, реснитчатых и секреторных клеток, гладкой мускулатуры [1-3]. В однорядном цилиндрическом трубном эпителии различают мерцательные (реснитчатые) и секреторные (бокаловидные) клетки. Базальные клетки способны дифференцироваться в мерцательные или секреторные. Под действием половых гормонов мерцательные и секреторные клетки могут превращаться друг в друга. В результате апоптоза и последующей дистрофии секреторные и мерцательные клетки превращаются в штифтообразные, которые затем «отшелушиваются» или подвергаются аутолизу. Секреторные клетки вырабатывают слизь, гликопротеины, электролиты, простагландины, факторы роста [4]. Эти вещества особенно активно выделяются в лютеиновую фазу менструального цикла и необходимы для капацитации сперматозоидов, жизнедеятельности яйцеклетки и эмбриона, регуляции процессов оплодотворения и ранних стадий развития эмбриона. На поверхности реснитчатого эпителия маточных труб плотность ресничек составляет около 1 млрд на 1 см. Главная функция этого огромного аппарата - создавать ток жидкости около поверхности клетки, скорость которого составляет 70-100 мкм/с [10]. С этим током жидкости происходит цилиарный транспорт - перенос яйцеклетки или эмбриона. Активности мерцательного эпителия отводится важная роль в обеспечении транспорта яйцеклетки и зародыша [3, 5-9]. Но размер яйцеклетки слишком велик, чтобы ее транспорт осуществлялся только за счет силы потока. Необходимыми условиями являются тесное взаимодействие и адгезивные контакты между рецепторами ресничек и клетками кумулюса яйцеклетки, которые делают транспорт более эффективным [5]. В осуществлении переноса по маточным трубам играет роль и перистальтика маточных труб. Точно оценить вклад гладкой мускулатуры трудно. Представляется, что мышечные сокращения обеспечивают захват яйцеклетки фимбриями, а также перемещение эмбриона, особенно по истмической части маточной трубы, где наибольшая (около 0,5 см) толщина мышечных стенок, меньше мерцательных и больше секреторных клеток, складки эндосальпинкса невысокие и лишены вторичного ветвления [4, 11]. Значение реснитчатого эпителия более выражено в ампуллярной части маточной трубы, где просвет ее значительно шире, толщина мышечной стенки в 5 раз меньше, чем в истмусе [4], концентрация мерцательных клеток наибольшая [12], складки слизистой оболочки высокие и ветвистые [11]. После захвата яйцеклетки фимбриями удержание ее в маточной трубе происходит при активном участии ресничек - наблюдается компактизация кумулюса яйцеклетки, он уменьшается в диаметре и становится способен на адгезивные контакты с эпителиоцитами [5]. В дальнейшем за продвижение яйцеклетки по маточной трубе отвечают главным образом реснички. Благодаря анатомическим и функциональным особенностям маточной трубы яйцеклетка может задержаться в ампуллярном отделе маточной трубы на 12 ч, чтобы дождаться оплодотворения. А затем эмбрион проходит расстояние 9-10 см именно за отведенные 4 сут и попадает в полость матки уже на стадии морулы [1]. Трудно представить, чтобы транспорт яйцеклетки и эмбриона по маточным трубам - процесс, расписанный по часам, зависел только от феномена «выдавливания» мышечными сокращениями, насколько тонкой должна быть их регуляция. Адгезивные контакты между клетками кумулюса и ресничками эпителиоцитов и ключевая роль мерцательного эпителия в процессе транспорта яйцеклетки объясняют точное соблюдение графика движения яйцеклетки и эмбриона. Частота биения ресничек будет прямо влиять на скорость продвижения эмбриона, а экспрессия молекул адгезии на поверхности клеток кумулюса «включает сцепление» и делает возможным перенос яйцеклетки. И если яйцеклетка попадает на участок эпителия, где реснички неподвижны или двигаются несогласованно, то ее продвижение останавливается, и беременность не наступает или развивается трубная беременность. Реснички на поверхности эпителиальных клеток представляют собой миниатюрные выросты плазматической мембраны с комплексом микротрубочек внутри. Толщина одной реснички составляет 0,25 мкм, а длина ее около 5 мкм [12]. На поверхности уже функционирующего эпителиоцита непрерывно протекают процессы цилиогенеза - вырастают новые реснички и отпадают старые [13, 14]. Каждая ресничка совершает движение, подобное удару хлыстом или гребку пловца. Цикл такого движения состоит из активной быстрой фазы, когда выпрямленная ресничка наклоняется и проталкивает вперед жидкость, и пассивной фазы, когда ресничка изгибается и возвращается в исходное положение, максимально прижимаясь к поверхности клетки [13, 15, 16]. В течение 1 с совершается 5-10 таких биений. В большинстве исследований частота биения ресничек в маточных трубах человека составляла 5-8 Гц [9, 17, 18]. In vivo [17] и in vitro [18] было показано, что в фимбриальном отделе частота биения ресничек на 0,5 Гц выше, чем в ампуллярном. Реснички работают координированно - двигательные циклы соседних ресничек едва заметно сдвинуты во времени. И если наблюдать под микроскопом за поверхностью мерцательного эпителия, то отчетливо видны ряды бегущих по его поверхности в одном направлении волн - это согласованно поднимаются и опускаются ряды ресничек. Координированное биение ресничек позволяет мерцательному эпителию с точной скоростью переносить эмбрион по направлению к полости матки. Циклические изменения в мерцательном трубном эпителии В течение менструального цикла в маточных трубах и в эндосальпинксе, как и во всех гормонально-зависимых органах, наблюдаются изменения. Структурные циклические изменения в эпителии наиболее выражены в ампуллярном и фимбриальном отделах, тогда как минимальны в перешейке и интрамуральном отделе [4, 19]. В первой половине менструального цикла поверхность эндосальпинкса ровная, гладкая. В фолликулярную фазу наиболее интенсивно идет образование новых мерцательных клеток, тогда как в лютеиновую фазу больше образуется секреторных [20]. Высота секреторных и мерцательных клеток в начале фолликулярной фазы обычно не превышает 20 мкм, к моменту овуляции достигает 30-35 мкм. На протяжении лютеиновой фазы менструального цикла высота мерцательных клеток снижается до 18-20 мкм в перешейке, тогда как в ампуллярном отделе не изменяется [20]. Образованию и росту мерцательных клеток и ресничек способствуют эстрогены. В фолликулярную фазу идет интенсивный цилиогенез. Незадолго до овуляции отмечается повышение числа реснитчатых клеток [21, 22]. В постменопаузе отмечается снижение как пролиферативной, так и функциональной активности мерцательного эпителия, появляются клетки с короткими ресничками или с небольшим их числом [23]. Регуляция цилиарного транспорта Цилиогенез - образование новых ресничек на поверхности эпителиоцита в маточной трубе, а также двигательная активность ресничек являются гормонально-зависимыми процессами и определяются числом рецепторов к половым гормонам в эпителиоцитах [14, 57]. Высокий уровень прогестерона может вызывать дисфункцию и даже «паралич» ресничек. При инкубации образцов трубного эпителия с добавлением гормонов в различных концентрациях показано, что при концентрации прогестерона 0,5 или 1 нг/мл частота биения ресничек снижалась на 63%, а при концентрации 2 нг/мл 50-70% ресничек были парализованы [24]. В другой работе инкубация с прогестероном (10 мкмоль/л) подавляла скорость биения ресничек на 40-50%, независимо от того, из какого отдела маточной трубы был получен биоптат [18]. Эффект прогестерона обратим, зависит от концентрации гормона и уменьшается при добавлении антагонистов прогестерона. Влияние эстрогенов на активность ресничек минимальна. Роль эстрогенов заключается в стимуляции дифференцировки клеток из секреторных в реснитчатые. Эстрадиол в концентрации 10 мкмоль/л не влиял на частоту биения ресничек, но уменьшал супрессорное влияние прогестерона [18]. Действие эстрадиола может проявляться в легком (на 4%) увеличении частоты биения [24]. При отсутствии эстрадиола мерцательные эпителиальные клетки становятся секреторными [25]. Тонкую местную регуляцию работы эпителия осуществляют паракринные системы. В частности, функционирование эпителия маточных труб зависит от выделяемых кумулюcом яйцеклетки факторов роста (инсулиноподобные факторы роста, фактор роста фибробластов, трансформирующий ростовой фактор), металлопротеиназы-I, урокиназы [2, 26]. Местные факторы, вызывающие активацию ресничек, приводят к преимущественной дифференцировке эпителиоцитов в мерцательные клетки [17]. Имеются сообщения о присутствии на эпителиальных клетках маточных труб рецепторов к ангиотензину II и о его стимулирующем влиянии на цилиарную активность [27]. Гонадотропины непосредственно не влияют на функционирование эпителия [24], но могут изменять его активность посредством эстрогенов и прогестерона. На фоне стимуляции овуляции под действием экзогенного хорионического гонадотропина наблюдается повышение числа секреторных клеток [28]. Факторы, нарушающие цилиарный транспорт Чаще всего повреждение эпителия маточных труб имеет инфекционную природу. Рассматриваются также механические повреждения маточных труб, наследственные нарушения структуры аксонемы, курение, медикаментозное и гормональное воздействие. Наиболее выраженное повреждение эпителия наблюдается при гонорее. Гонококковая инфекция вызывает значительные нарушения цилиарной активности и слущивание реснитчатых клеток. Этот процесс вызван липидом А липополисахарида гонококка, фрагментами пептидогликана и такими цитокинами, как фактор некроза опухоли-a [31, 32]. Chlamydia trachomatis обладает способностью персистировать в клетках цилиндрического эпителия. Хламидии не вызывают тотальную гибель эпителия, а поражают лишь ту клетку, которую инфицируют. Это приводит к потере ресничек и нарушению межклеточных контактов. А через 24 ч после инфицирования наблюдаются разрушение эпителиоцита и высвобождение хламидийных телец [33]. Также отмечалось троекратное снижение частоты биения ресничек эпителия маточных труб, пораженных хламидийной инфекцией [34]. Примечательно, что у женщин с антителами к хламидиям серотипов С и Е (без D) отмечается более низкая частота биения ресничек, чем у женщин с хламидийной инфекцией, вызванной другими серотипами [35]. В культуре зараженных Mycoplasma hominis эпителиоцитов маточных труб наблюдались замедление и дискоординация биения ресничек, а также появление клеток с отсутствием активности ресничек [36, 37]. Выраженное повреждение эпителиоцитов и ресничек имеет место при туберкулезной инфекции маточных труб [38]. Экстракт Bacteroides ureolyticus вызывает паралич ресничек трубного эпителия [39]. У женщин, использовавших внутриматочные контрацептивные средства, число мерцательных клеток уменьшается в 2 раза [40]. Стерилизация путем перевязки маточных труб вызывает дистрофию эпителия с потерей цилиарной активности в пределах 0,5 см от наложенной лигатуры. На расстоянии 1 см эпителий функционирует нормально [41]. Катетер, диаметром 0,66 мм, использующийся для переноса эмбрионов в маточные трубы, не вызывает повреждения эпителия [42]. При синдроме Картагенера, синдроме первичной цилиарной дискинезии - генетически обусловленном заболевании, нарушается структура аксонемы. Это могут быть как потери центральных микротрубочек, так и нарушения в работе белков - регуляторов движения микротрубочек. Нарушается, а иногда и полностью «парализуется» работа значительной части ресничек мерцательного эпителия, независимо от того, в каком органе находятся клетки. Наблюдается сочетание хронического синусита, бронхоэктатической болезни и бесплодия. Встречается полная инверсия внутренних органов [7, 8, 43, 44, 56, 59, 60]. Эпидемиологические исследования показывают, что у курящих женщин чаще встречаются бесплодие, непроходимость маточных труб и внематочные беременности. Все тестируемые компоненты табачного дыма (никотин, акролеин, формальдегид, фенол, ацетальдегид, цианистый калий) дозозависимо и обратимо снижали частоту биения ресничек в 3-50 раз [45-47]. Также наблюдалось зависимое от дозы снижение скорости продвижения яйцеклетки по маточной трубе даже при сохраненной частоте биения ресничек, что объясняется нарушением адгезивных контактов между ресничками и кумулюсом. Гормональные нарушения или медикаментозная терапия, сопровождающиеся высоким уровнем прогестерона и приводящие к замедлению биения ресничек или их полному «параличу», могут быть одной из причин нарушения транспорта яйцеклетки по маточной трубе [18, 24]. Негативно на работе мерцательного аппарата сказываются гипоэстрогенные состояния [24, 26]. Стимуляция овуляции у крольчих кломифеном в обычных дозах не вызывала значительных изменений в состоянии маточных труб, тогда как доза больше 5 мг/кг вызывала дискоординированное биение ресничек [29]. В эксперименте на коровах в результате стимуляции овуляции происходило увеличение числа мерцательных клеток, а также возрастало число и активность ресничек на их поверхности [30]. На фоне стимуляции овуляции наблюдается качественное и количественное повышение активности мерцательного эпителия или дискоординация биения ресничек. Не исключено, что это может приводить к нарушению транспорта яйцеклетки (преждевременному попаданию эмбриона в полость матки или его задержке в маточной трубе). В эксперименте показано, что антагонист кальция нифедипин вызывает дискоординацию сокращений гладкой мускулатуры маточной трубы, а также нерегулярное биение ресничек, влияя на их энергетическое митохондриальное обеспечение [48]. Поражение цилиарного транспорта может иметь разную природу: гибель эпителиальных клеток, потеря ресничек, снижение частоты биения ресничек, дискоординация биения ресничек, нарушение адгезивной способности к клеткам кумулюса. Все эти нарушения могут прямо или косвенно быть причиной бесплодия. Цилиарный транспорт и воспалительные заболевания придатков матки Кроме функции переноса эмбриона, цилиарный транспорт играет роль «дворника» эндосальпинкса. Благодаря биению ресничек маточная труба очищается от секрета, попавших в нее микроорганизмов, погибших сперматозоидов и менструальных выделений. Строение и принципы работы ресничек цилиндрического эпителия разных органов одинаковы, будь то респираторный тракт, маточные трубы или носовые пазухи [7, 8, 12, 15]. Тогда как в пульмонологии и отоларингологии нарушению мукоцилиарного транспорта придается большое значение в развитии хронических воспалительных заболеваний, в гинекологии до настоящего времени этот феномен практически не рассматривается. Концентрация бактерий во влагалище составляет 7-8 lg микробных тел/мл, при бактериальном вагинозе повышается еще на 4-5 порядков, тогда как верхние отделы репродуктивной системы считаются стерильными. Возможность проникновения туда микроорганизмов самостоятельно или с участием сперматозоидов не вызывает сомнения. Адекватное очищение матки и маточных труб обеспечивает, помимо местных факторов иммунитета, мукоцилиарный транспорт. Как правило, инициаторами воспалительного процесса в органах малого таза являются специфические возбудители (гонококки, хламидии, микоплазмы). Эти микроорганизмы обладают способностью вызывать значительные функциональные нарушения в трубном эпителии. Особенно это относится к гонорейной и хламидийной инфекциям. Эпителий может поражать энтеровирусная инфекция. Слущивание эпителиальных клеток, снижение частоты биения ресничек, дискоординация их активности создают условия для персистирования и размножения в маточных трубах условно-патогенных аэробов и анаэробов, которые сами обладают способностью вызывать нарушения в работе ресничек и могут длительно поддерживать воспаление даже после излечения специфической инфекции. Эта концепция хорошо объясняет существующие в настоящее время представления о смешанной аэробно-анаэробной этиологии воспалительных процессов органов малого таза и ведущей роли облигатно-анаэробных бактерий [49]. В результате воспалительной реакции в маточных трубах наблюдаются явления отека, нарушения тканевого обмена, нарушения микроциркуляции [50]. Это только усугубляет дисфункцию эпителия. У 62 женщин с трубно-перитонеальным бесплодием была произведена биопсия маточных труб. Световая, электронная микроскопия и лазерная спектроскопия показали, что частота биения ресничек была значительно ниже в тканях с отеком и полнокровием, чем в тканях без отека и полнокровия [35]. Неадекватный мукоцилиарный клиренс слизистой маточных труб приводит к проникновению микроорганизмов в ампуллярную и фимбриальную части маточной трубы и далее - в брюшину малого таза. Ответная воспалительная реакция брюшины в виде образования спаек препятствует дальнейшему распространению микроорганизмов и развитию пельвиоперитонита, хотя в ряде случаев это происходит. При выраженном микробном размножении в маточных трубах иммунная система реагирует не только экссудативным, но и инфильтративным воспалением. Начинаются процессы склероза и рубцевания, роль которых в развитии окклюзии и непроходимости труб хорошо известна. Чаще всего облитерация просвета маточной трубы происходит в истмической части трубы. Это связано с более узким просветом в данном отделе, а также меньшим количеством мерцательных клеток в истмическом отделе, чем в ампуллярном. Поражение эпителия в истмусе скорее приведет к нарушению цилиарного транспорта. Цилиарный транспорт и трубная беременность Трубная беременность развивается из-за замедленного продвижения эмбриона по маточной трубе или его задержки в ней. Основной причиной этого, традиционно, считается дисфункция нервно-мышечного аппарата маточных труб, которая развивается при воспалительных заболеваниях и гормональных нарушениях. В то же время практически все состояния, способствующие внематочной беременности, являются факторами, которые связаны с замедлением, дискоординацией биения ресничек, либо с повреждением трубного эпителия. К ним относятся воспалительные заболевания органов малого таза, операции на маточных трубах, экстракорпоральное оплодотворение, эндометриоз, эндокринные нарушения, применение внутриматочных контрацептивов, генитальный инфантилизм. При попадании эмбриона на участок эндосальпинкса с недостаточной активностью ресничек происходит незапланированная задержка его продвижения. По мере созревания трофобласта выделяются протеолитические ферменты и происходит имплантация в стенку трубы, развивается трубная беременность. Если же имплантации не происходит, то зародыш гибнет на ранней стадии развития [58]. При трубной беременности сперматозоид к яйцеклетке проникнуть может. Значит труба все-таки проходима. Но вот эмбриону попасть в полость матки не суждено. Можно предположить, что дело в маленьких размерах сперматозоида и в больших - яйцеклетки. Но нельзя забывать еще об одном существенном различии между ними. Если сперматозоид подвижен и в посторонней помощи не нуждается, то яйцеклетка тяжела и неповоротлива. Реснички трубного эпителия способны бережно перенести ее в полость матки. Если же они поражены на одном из участков трубы из-за воспаления, то развивающийся зародыш остается на месте. Часто трубная беременность возникает сначала в одной, а через некоторый период и в другой трубе, потому что повреждающие трубный эпителий инфекционные и другие факторы действуют неизбирательно и поражают эпителий обеих труб. В ряде работ подтверждается роль патологии мерцательного эпителия в развитии трубной беременности. При изучении эпителия фимбриального отдела маточных труб у женщин с трубной или маточной беременностью установлено, что при внематочной беременности имеет место атрофия эпителия, проявляющаяся потерей ресничек, снижением высоты реснитчатых клеток и нарушением распределения кальция между митохондриями и цитоплазмой [51]. В другом исследовании при трубной беременности по сравнению с маточной отмечалось существенное снижение числа мерцательных клеток [52]. Правда, тонких ультраструктурных различий в строении ресничек при внематочной и маточной беременности при сканирующей электронной микроскопии поверхности эпителия маточных труб не обнаружено [53]. Основную роль играют характер биения ресничек и молекулы адгезии. Мы нисколько не умаляем роль нарушения перистальтики маточных труб при развитии трубной беременности. Но выше мы уже высказывали сомнение относительно значимости феноменов «выдавливания» и считаем, что работа мерцательного эпителия играет решающую роль. Цилиарный транспорт и эндометриоз Хотя патогенез эндометриоза изучается более ста лет, многие вопросы еще неясны. Наиболее обоснованной в настоящее время считается имплантационная (трансплантационная) теория, согласно которой эндометриоз возникает в результате имплантации и роста ретроградно распространяющихся клеток эндометрия. Имплантационная теория предполагает соблюдение трех условий: ретроградный ток менструальной крови, наличие в ней жизнеспособных клеток и их прикрепление вне полости матки [10]. Определяющими для развития эндометриоза будут условия, в которых окажутся клетки эндометрия (молекулы адгезии, реакция иммунокомпетентных клеток и свертывающей системы крови). Но факт переноса клеток через маточные трубы в имплантационной теории является обязательным условием. Менструальная кровь практически всегда попадает в маточную трубу и приблизительно в 90% случаев проникает в брюшную полость, у женщин с эндометриозом во время менструаций клетки эндометрия чаще обнаруживаются в брюшной полости [10, 54]. Прохождению менструальных выделений через маточную трубу препятствуют как перистальтические сокращения мышечной оболочки, так и мукоцилиарный транспорт на поверхности эпителия. Вероятно, что при нарушении мукоцилиарного транспорта, при неадекватной активности ресничек или недостаточном числе мерцательных клеток создаются условия, способствующие ретроградному перемещению большего количества менструальных выделений по маточной трубе. Истинную роль нарушений цилиарного транспорта в развитии эндометриоза еще предстоит выяснить. Не исключено, что бессимптомно протекающий эндометриоз и ретроградные менструации физиологичны [54]. Если в итоге окажется, что ретроградных менструаций быть не должно, то нарушение цилиарного транспорта займет значимое место в патогенезе наружного эндометриоза и эндометриозассоциированного спаечного процесса в малом тазу. Если же подтвердится теория о физиологичности ретроградных менструаций, то нарушения цилиарного транспорта могут быть причиной попадания в брюшную полость слишком большого количества менструальной крови и развития ответного спаечного процесса в полости малого таза. Не исключается, что эндометриоз является только кофактором бесплодия, а не его непосредственной причиной. В этом свете причиной трубного бесплодия на фоне эндометриоза может быть нарушение цилиарного транспорта. Цилиарный транспорт и трубное бесплодие О том, как нарушение цилиарного транспорта может приводить к бесплодию, мы уже упоминали, когда говорили о воспалительных заболеваниях маточных труб, спаечном процессе в области малого таза, эндометриозе, трубной беременности. Все эти состояния хорошо известны как причина трубно-перитонеального бесплодия у женщин. Центральную роль в развитии трубного бесплодия играют нарушения цилиарного транспорта (см. схему). Но возможно существование еще одной формы трубного бесплодия. На наш взгляд, некоторые формы трубного бесплодия могут протекать с проходимостью маточной трубы для контрастного вещества при гистеросальпингографии или красителя при лапароскопии, т.е. диагноз трубного фактора бесплодия клинически исключается. Но в действительности на фоне воспалительного процесса, рубцовых изменений или других причин могут существовать дефект эпителия, нарушение цилиарного транспорта и не анатомическая, а функциональная непроходимость, когда задержка продвижения яйцеклетки по маточной трубе будет приводить к прерыванию беременности еще до имплантации. Центральная роль цилиарной дискинезии мерцательных клеток эндосальпинкса в патогенезе трубного бесплодия.
Доля мерцательных эпителиоцитов у бесплодных и фертильных женщин может не различаться, гораздо важнее их функциональное состояние - частота и координированность биения ресничек, адекватное кровоснабжение эндосальпинкса [56]. С развитием методов исследования будет уделяться больше внимания изучению состояния реснитчатого эпителия маточных труб (равно как и изучению работы жгутиков сперматозоидов), будут разработаы критерии нормы и патологии, например, по таким параметрам, как доля мерцательных клеток, частота биения ресничек, скоординированность их биения, число реcничек на эпителиальной клетке. В будущем станет возможным диагностировать состояние цилиарной дискинезии с помощью уже разработанного метода тубоскопии с микроскопией реснитчатого эпителия [17]. Представляется, что у многих пациенток, страдающих хроническими воспалительными заболеваниями придатков, внематочными беременностями, наружным эндометриозом или некоторыми гормональными нарушениями, в диагнозе будет присутствовать цилиарная дискинезия маточных труб. С.М. Белобородов Научный центр акушерства, гинекологии и перинатологии РАМН,Москва Литература 1. Buhi W.C., Alvarez I.M., Kouba A.J. Oviductal regulation of fertilization and early embryonic development. J Reprod Fertil Suppl 1997; 52: 285-300. 2. Einspanier R., Lauer B., Gabler C., Kamhuber M., Schams D. Egg-cumulus-oviduct interactions and fertilization. Adv Exp Med Biol 1997; 424: 279-89. 3. Gordons D. Tubal infertility. New York 1996; 145. 4. Железнов Б.И. Репродуктивные органы репродуктивной системы. Руководство по эндокринной гинекологии. Под ред. Е.М. Вихляевой. М: МИА 1997; 768. 5. Talbot P., Geiske C., Knoll M. Oocyte pickup by the mammalian oviduct. Mol Biol Cell 1999; 10(1): 5-8. 6. Afzelius B.A. Role of cilia in human health. Cell Motil Cytoskeleton 1995; 32(2): 95-7. 7. McComb P., Langley L., Villalon M., Verdugo P. The oviductal cilia and Kartagener's syndrome. Fertil Steril 1986; 46(3): 412-6. 8. Halbert S.A., Patton D.L., Zarutskie P.W., Soules M.R. Function and structure of cilia in the fallopian tube of an infertile woman with Kartagener's syndrome. Hum Reprod 1997; 12(1): 55-8. 9. DiCarlantonio G., Shaoulian R., Knoll M., Magers T., Talbot P. Analysis of ciliary beat frequencies in hamster oviductal explants. J Exp Zool 1995; 272: 142-152. 10. Linden P.J.Q. Endometriosis pathogenesis teories. Hum Reprod 1996; 11(3): 59-72. 11. Кондриков Н.И. Гистофизиологические особенности маточных труб. Акуш и гинекол 1989; 2: 38-44. 12. Dirksen E.R., Zeira M. Microtubule sliding in cilia of the rabbit trachea and oviduct. Cell Motil 1981; 1(2): 247-60. 13. Hagiwara H., Aoki T., Ohwada N., Fujimoto T. Development of striated rootlets during ciliogenesis in the human oviduct epithelium. Cell Tissue Res 1997; 290(1): 39-42. 14. Cornier E., Chatelet F., Grenier J., Valade S., Salat-Baroux J., Roland J. Epithelium cilie de la trompe de Fallope chez la femme. Etude ultrastructurale. J Gynecol Obstet Biol Reprod 1980; 9(5): 505-11. 15. Албертс Б., Брей Д., Лбюис Дж., Рэфф М., Робертс К., Уотсон Дж. Молекулярная биология клетки. М: Мир 1994; 1: 530. 16. Hagiwara H., Aoki T., Fujimoto T. Ultrastructural observation on "transitional tubules" in human oviductal ciliogenic cells. J Anat 1997; 191(2): 285-90. 17. Paltieli Y., Weichselbaum A., Hoffman N., Eibschitz I., Kam Z. Laser scattering instrument for real time in vivo measurement of ciliary activity in human fallopian tubes. Hum Reprod 1995; 10(7): 1638-41. 18. Mahmood T., Saridogan E., Smutna S., Djahanbakhch O., Habib A.M. The effect of ovarian steroids on epithelial ciliary beat frequency in the human Fallopian tube. Hum Reprod 1998; 13(11): 2991-4. 19. Abe H., Onodera M., Sugawara S. Scanning electron microscopy of goat oviductal epithelial cells at the follicular and luteal phases of the oestrus cycle. J Anat 1993; 183: 415-21. 20. Abe H., Oikawa T. Observations by scanning electron microscopy of oviductal epithelial cells from cows at follicular and luteal phases. Anat Rec 1993; 235(3): 399-410. 21. Hagiwara H., Shibasaki S., Ohwada N. Ciliogenesis in the human oviduct epithelium during the normal menstrual cycle. J Electron Microsc 1992; 41 (5): 321-9. 22. Shirley B., Reeder R.L. Cyclic changes in the ampulla of the rat oviduct. J Exp Zool 1996; 276(2): 164-73. 23. Frappart L., Berger G., Bethouart M., Palayer C., Bremond A., Rochet Y., Feroldi J. L'epithelium tubaire. Microscopie electronique a balayage au cours du cycle menstruel, de la grossesse et de la menopause. J Gynecol Obstet Biol Reprod (Paris) 1980; 9(3): 307-13. 24. Paltieli Y., Eibschitz I., Ziskind G., Ohel G., Silbermann M., Weichselbaum A. High progesterone levels and ciliary dysfunction-a possible cause of ectopic pregnancy. J Assist Reprod Genet 2000; 17(2): 103-6. 25. Mahmood T. et al. The effect of ovarian steroids on epitelial ciliary beat frequency in the human fallopian tube. Hum Reprod 1995; 10 (7): 1638-41. 26. Einspanier R., Gabler C., Bieser B., Einspanier A., Berisha B., Kosmann M., Wollenhaupt K., Schams D. Growth factors and extracellular matrix proteins in interactions of cumulus-oocyte complex, spermatozoa and oviduct. J Reprod Fertil Suppl 1999; 54: 359-65. 27. Saridogan E., Djahanbakhch O., Puddefoot J.R., Demetroulis C., Collingwood K., Mehta J.G., Vinson G.P. Angiotensin II receptors and angiotensin II stimulation of ciliary activity in human fallopian tube. J Clin Endocrinol Metab 1996; 81(7): 2719-25. 28. Bondi A.M., Gabrielli M.G., Marchetti L., Materazzi G., Menghi G. Cytomorphological changes in the rabbit oviductal epithelium after human chorionic gonadotropin treatment. Histol Histopathol 1997; 12(1): 135-46. 29. Birkenfeld A., Weber-Benndorf M. Effect of clomiphene on oviductal and uterine mucosa functional morphology. Ann NY Acad Sci 1985; 442: 153-67. 30. Abughrien B.M., Dore M.A. Ciliogenesis in the uterine tube of control and superovulated heifers. Cells Tissues Organs 2000; 166(4): 338-48. 31. Stephens D.S., McGee Z.A., Cooper M.D. Cytopathic effects of the pathogenic Neisseria. Studies using human fallopian tube and human nasopharyngeal organ cultures. Antonie Van Leeuwenhoek 1987; 53(6): 575-84. 32. McGee Z.A., Jensen R.L., Clemens C.M., Taylor-Robinson D., Johnson A.P., Gregg C.R. Gonococcal infection of human fallopian tube mucosa in organ culture: relationship of mucosal tissue TNF-alpha concentration to sloughing of ciliated cells. Sex Transm Dis 1999; 26(3): 160-5. 33. Cooper M.D., Rapp J., Jeffery-Wiseman C., Barnes R.C., Stephens D.S. Chlamydia trachomatis infection of human fallopian tube organ cultures. J Gen Microbiol 1990; 136: 1109-15. 34. Patton D.L., Moore D.E., Spadoni L.R., Soules M.R., Halbert S.A., Wang S.P. A comparison of the fallopian tube's response to overt and silent salpingitis. Obstet Gynecol 1989; 73(4): 622-30. 35. Leng Z., Moore D.E., Mueller B.A., Critchlow C.W., Patton D.L., Halbert S.A., Wang S.P. Characterization of ciliary activity in distal Fallopian tube biopsies of women with obstructive tubal infertility. Hum Reprod 1998; 13(11): 3121-7. 36. Baldetorp B., Mardh P.A., Westrom L. Studies on ciliated epithelia of the human genital tract. Mucociliary activity in organ cultures of human fallopian tubes challenged with Mycoplasma hominis. Sex Transm Dis 1983; 10: 363-5. 37. Swenson C.E., Banks J.P., Schachter J. Organ culture studies with Mycoplasma hominis. Sex Transm Dis 1983; 10: 355-8. 38. Biolatti B., Pau S., Galloni M. The epithelial pathology of bovine genital tuberculosis. J Comp Pathol 1989; 100(2): 137-44. 39. Fontaine E.A., Clark J.B., Abeck D., Taylor-Robinson D. The effect of a toxin from on the mucosal epithelium of human and bovine oviducts. Br J Exp Pathol 1988; 69(5): 631-8. 40. Wollen A.L., Flood P.R., Sandvei R., Steier J.A. Morphological changes in tubal mucosa associated with the use of intrauterine contraceptive devices. Br J Obstet Gynaecol 1984; 91(11): 1123-8. 41. Li J., Chen X., Zhou J. Ultrastructural study on the epithelium of ligated fallopian tubes in women of reproductive age. Anat Anz 1996; 178(4): 317-20. 42. Kitamura S., Miyazaki T., Iwata S., Akaboshi K., Osawa Y., Yoshimura Y. Ultrastructural evaluation following catheterization of the fallopian tube with a hysteroscopic catheter. J Assist Reprod Genet 1998; 15(7): 411-7. 43. Бочков Н.П. Клиническая генетика. М: Медицина 1997; 288. 44. Roperto F., Saviano R., Guarino G. Atypical basal bodies in a cat with immotile-cilia syndrom. J Submicrosc Cytol Pathol 1994; 26(4): 565-7. 45. Talbot P., DiCarlantonio G., Knoll M., Gomez C. Identification of cigarette smoke components that alter functioning of hamster oviducts in vitro. Biol Reprod 1998; 58(4): 1047-53. 46. Knoll M., Talbot P. Cigarette smoke inhibits oocyte cumulus complex pick-up by the oviduct in vitro independent of ciliary beat frequency. Reprod Toxicol 1998; 12(1): 57-68. 47. Knoll M., Shaoulian R., Magers T., Talbot P. Ciliary beat frequency of hamster oviducts is decreased in vitro by exposure to solutions of mainstream and sidestream cigarette smoke. Biol Reprod 1995; 53(1): 29-37. 48. Yang L., Xu Y.X. Effect of nifedipine on contractive motility and ciliary ultrastructure of rabbit fallopian tube. Chung Kuo Yao Li 1993; 14(5): 450-2. 49. Кулаков В.И., Воропаева С.Д., Анкирская А.С. Облигатно-анаэробные микроорганизмы при акушерско-гинекологической патологии. Вестник РАМН 1996; 2: 26-29. 50. Стрижаков А.Н., Подзолкова Н.М. Гнойные воспалительные заболевания придатков матки. M: Медицина 1996; 255. 51. Peretz B.A., Lindenbaum E.S., Beach D. Ectopic pregnancy effects on the ipsilateral fallopian tube epithelium-an ultrastructural study. Eur J Obstet Gynecol Reprod Biol 1984; 17(1): 19-28. 52. Vasquez G., Winston R.M., Brosens I.A. Tubal mucosa and ectopic pregnancy. Br J Obstet Gynaecol 1983; 90(5): 468-74. 53. Samberg I., Degani S., Zilberman A., Eibschitz I., Nir I., Scharf M. Scanning electron microscopy of human Fallopian tube in ectopiс pregnancy. Gynecol Obstet Invest 1983; 16(2): 65-75. 54. Thomas E.J. К проблеме бессимптомно протекающего эндометриоза. Hum Reprod 1996; 11(3): 113-121. 55. Lurie M., Tur-Kaspa I., Weill S., Katz I., Rabinovici J., Goldenberg S. Ciliary ultrastructure of respiratory and fallopian tube epithelium in a sterile woman with Kartagener's syndrome. A quantitative estimation. Chest 1989; 95(3): 578-81. 56. Marchini M., Baglioni A., Zamberletti D., Vercellini P., Motta T., Fedele L. Ultrastructure of endosalpingeal biopsies in infertile patients: correlation with reproductive success. Int J Gynaecol Obstet 1988; 27(2): 239-45. 57. Comer M.T., Leese H.J. Induction of a differentiated ciliated cell phenotype in primary cultures of tube epithelium. Hum Reprod 1998; 13(11): 3114-20. 58. Buhi W.C., Alvarez I.M., Kouba A. Oviductal regulation of fertilization and early embryonic development. J Reprod Fertil Suppl 1997; 52: 285-300. 59. Roperto F., Mingozzi C., Volpe P., Bruno F. Atypical cilia in the oviductal epithelium of healthy dogs. J Submicrosc Cytol Pathol 1998; 30(4): 581-3. 60. Roperto F., De Vico G., Maiolino P. Atypical cilia in the oviductal epithelium of healthy cats. J Submicrosc Cytol Pathol 1995; 27(2): 267-70. Уважаемые коллеги! На Русском Медицинском Сервере в сети Internet работает профессиональный форум по акушерству и гинекологии, где проводятся дискуссии по проблемам нарушений менструального цикла, бесплодного брака, половых инфекций, гормонотерапии, контрацепции и т. д. На форуме имеется раздел, где есть возможность обсуждения сложных клинических случаев, обмена опытом и мнениями. Приглашаем Вас присоединиться к дискуссиям по адресу: http://www.rusmedserv.com/cgi-bin/forum/UltraBoard.cgi Администрация Русского Медицинского Сервера Жена мужу: - Пока не съешь Васькин контрасекс, в санаторий не поедешь.
|